19.08.2019
Da doch einige Behauptungen und Annahmen durch die Presse rauschen von wegen schädlich oder nicht schädlich vom Mobilfunk, habe ich mich dem Thema über die wissenschaftliche Literatur genähert. Hierfür habe ich die Veröffentlichungen von Januar-August 2019 herausgesucht und gelesen. Dabei habe ich sieben wissenschaftliche Veröffentlichungen gefunden die relevant wären und die ich hier kurz besprechen möchte.
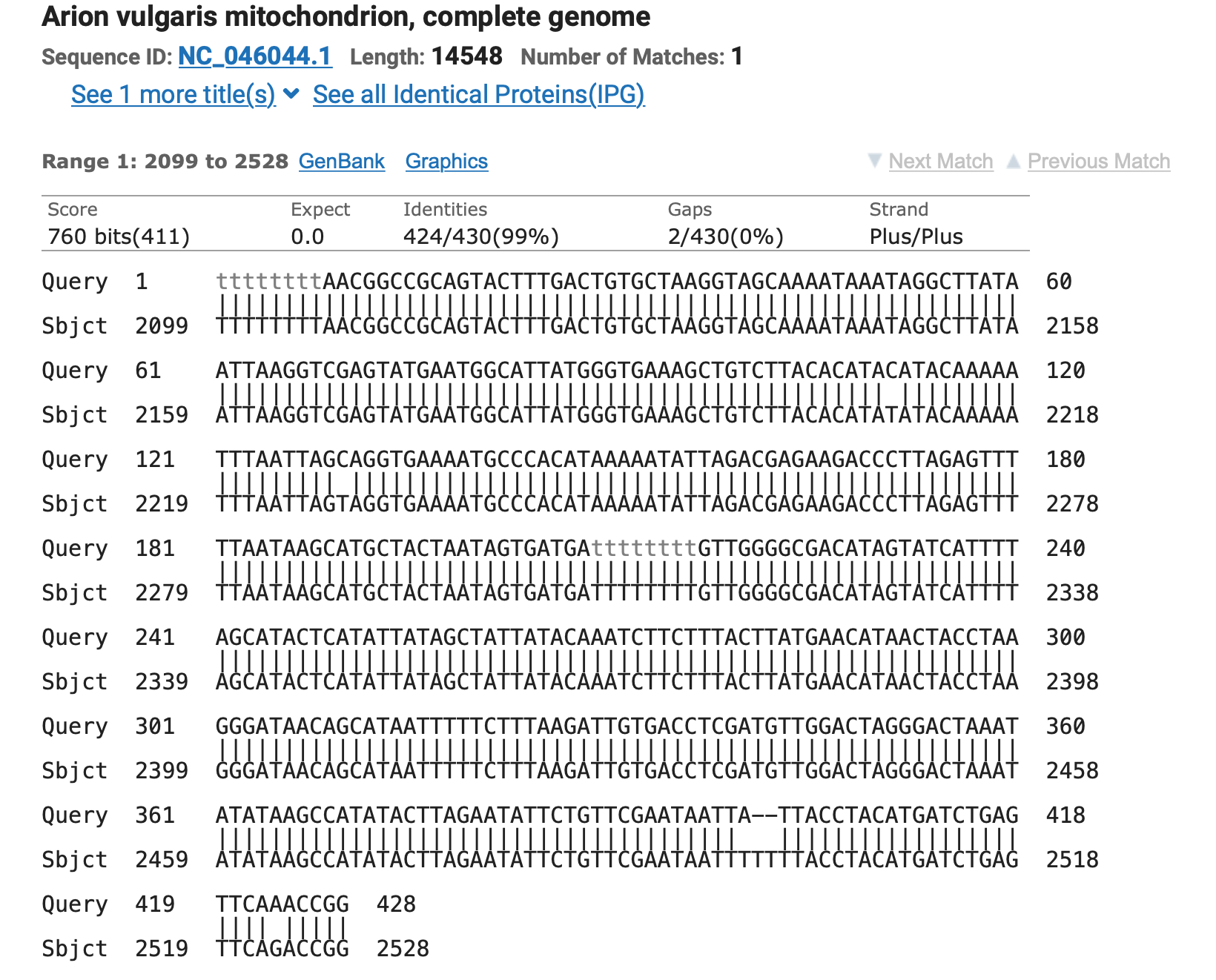
Die erste Studie hat mich selber überrascht, es ist eine Studie aus Schweden bei der bei über 24.000 Menschen Kopfschmerzen, Tinnitus und Schwerhörigkeit in Bezug auf die Nutzung von Mobiltelefonen untersucht wurde (1).
Resultat der schwedischen Studie: Personen, die am häufigsten Mobiltelefone verwendeten, berichteten etwas häufiger über wöchentliche Kopfschmerzen als andere Benutzer. Dieser Befund verschwand jedoch nach Anpassung der Störfaktoren weitgehend und hing nicht mit der Anrufdauer bei GSM mit höheren HF-Frequenzen zusammen. Tinnitus und Hörverlust waren nicht mit der Anrufdauer verbunden.
In der zweiten Studie wurden die kognitive Fähigkeiten bei Kinder mit Handy untersucht (2).
Die Autoren sagen, dass frühere epidemiologische Studien zu gesundheitlichen Auswirkungen der Strahlenexposition von Mobiltelefonen zu inkonsistenten Ergebnissen geführt hätten.
Es wurden 619 Schüler zu Studienbeginn rekrutiert und Längsschnittdaten für 412 Schüler aus einer repräsentativen Stichprobe von 36 staatlichen, privaten und katholischen Schulen erhoben.
Es gab kaum Anhaltspunkte für einen signifikanten Zusammenhang zwischen der Nutzung von Mobiltelefonen und der kognitiven Leistung von Schülern (2).
Eine dritte Studie bietet einen ganz guten Überblick über die elektromagnetische Felder in verschiedenen Umgebungen und erläutert diese (3).
Bei einer vierten Studie wird es wieder spannender, weil es geht um Tumore des Gehirns und der Speicheldrüse und Verwendung von Mobiltelefonen (4). Hier handelt es sich um eine sogenannte “Meta-Studie”. Die Veröffentlichung kombiniert eine Vielzahl von Studien in einer Zusammenschau. Hier die Ergebnisse der Studie:
Zusammenfassend lässt sich sagen, dass die aktuellen Erkenntnisse aus allen verfügbaren Studien, einschließlich in-vitro-, in-vivo– und epidemiologischen Studien, keinen Zusammenhang zwischen dem Mobiltelefon Konsum und Tumoren erkennen lassen, die sich aus den am stärksten exponierten Organen und Geweben entwickeln. Angesichts des großen Forschungsaufwands zu diesem Thema ist davon auszugehen, dass potenziell nicht erkannte Risiken aus individueller Sicht gering sind und lange Latenzzeiten (> 15 Jahre), seltene Gehirntumorsubtypen und Mobiltelefon-Konsum im Kindesalter betreffen können. Um diesen geringen Risiken zu begegnen, ist eine qualitativ hochwertige Forschung mit genauer Expositionsabschätzung erforderlich, wobei zu berücksichtigen ist, dass die Dauer des MP-Anrufs allein die HF-EMF-Exposition des Gehirns nicht angemessen widerspiegeln wird.
Bei der 5. Studie geht es auch wieder um Krebs, insbesondere um diverse Hirntumore (5).
In einer populationsbasierten ökologischen Studie wurden die Trends von Hirntumoren in den Zeiträumen 1982–1992, 1993–2002 und 2003–2013 untersucht. Die Schlussfolgerungen der australischen Wissenschaftler ergaben keine Zunahme von Gehirntumor-Histologietypen oder Gliomlokalisationen, die auf Mobiltelefone zurückgeführt werden können.
Dann in der 6. Studie ergaben sich dann doch Auswirkungen von Mobilfunkmasten neben Schulen auf die Konzentrationsfähigkeit von Schulkindern (6). Allerdings gibt der Autor – ein Professor Sultan Ayoub Meo von der King Saud Universität in Riad – zu, dass seine Statistik mangelhaft sei. Er belegt seine Aussagen dann aber mit einigen hochkarätigen Veröffentlichung unter anderem einer vom Max-Planck-Institut für neurologische Forschung (7).
Bei dieser 13-seitigen Veröffentlichung scheint es sich um sowas wie ein Standardwerk zu handeln. Es wird sowohl die Blut-Hirn Schranke genau behandelt wie auch alle anderen physiologischen und neurologischen Zusammenhänge. Ich will hier nur kurz die Schlussfolgerungen des Max-Planck-Institut für neurologische Forschung aus (7) zusammenfassen:
Gegenwärtig gibt es kaum Hinweise darauf, dass eine gepulste oder kontinuierliche Mikrowellenexposition bei Leistung und Frequenzen im Zusammenhang mit der Mobilkommunikation die Funktionsfähigkeit und strukturelle Integrität des Gehirns beeinträchtigen könnte. Unter experimentellen Bedingungen konnten die meisten der bisher gemeldeten positiven Ergebnisse auf thermische Effekte zurückgeführt werden. Es ist unwahrscheinlich, dass solche Effekte bei regelmäßiger Verwendung von Mobiltelefonen auftreten, da die emittierte Gesamtleistung viel zu niedrig ist, um die Körpertemperatur zu erhöhen, und weil lokale Erhöhungen der Gehirntemperatur, falls vorhanden, durch die thermostabilisierende Wirkung des zirkulierenden Blutes verhindert würden.
Andere biologische Störungen, die unter Versuchsbedingungen beobachtet werden, können mit verfahrensbedingten Nebenwirkungen wie Immobilisierungsstress zusammenhängen, unter denen nicht betäubte Tiere leiden. Versuchsanordnungen sollten daher so konzipiert werden, dass die tatsächlichen Expositionsbedingungen für menschliche Benutzer von Mobilkommunikationssystemen nachgebildet werden.
Ebenso sind Wechselwirkungen mit menschlichem Verhalten oder neurologischen Erkrankungen bestenfalls minimal. Insbesondere konnte kein signifikanter Zusammenhang mit der Inzidenz oder dem Wachstum von Hirntumoren unter experimentellen oder klinischen Bedingungen hergestellt werden. Negative Daten schließen die Möglichkeit geringfügiger mikrowelleninduzierter biologischer Wirkungen auf das Gehirn nicht aus. Je geringer diese Nebenwirkungen sind, desto geringer ist ihr Beitrag zum kombinierten Gesundheitsrisiko, das andere Umwelteinflüsse auf das Gehirn ausüben können. Diese Schlussfolgerung steht möglicherweise im Widerspruch zu den offensichtlichen, aber wenig untersuchten Stressreaktionen, die viele Personen erfahren, wenn sie sich über rücksichtslose Benutzer von Mobiltelefonen ärgern. Beträchtliche Gesundheitsrisiken ergeben sich auch aus der hohen Anzahl von Verkehrsunfällen, die durch Mobiltelefonie während der Fahrt verursacht werden. Weitere Forschungen sollten sich daher auf diese indirekten Auswirkungen der Mobiltelefonie konzentrieren, die sich letztendlich als schwerwiegendere Belastung für die Gesundheitssysteme herausstellen können als die vermutlich vernachlässigbaren biologischen Störungen.
Professor Sultan Ayoub Meo will seine Studie mit einer weiteren wichtigen amerikanischen Veröffentlichung belegen. In dieser Studie wurde schon 2004 gezeigt, dass die aktuellen Beweise für einen ursächlichen Zusammenhang zwischen Krebs und Exposition gegenüber Hochfrequenzenergie schwach und nicht überzeugend sind (8).
Fazit aus der Recherche von Januar-August 2019
Die einzige wissenschaftliche Veröffentlichung von 2019 die den Mobilfunk kritisch sieht, hat nicht nur keinerlei Beweise für die Behauptung, sondern in den zitierten Journals steht auch das Gegenteil der Behauptung.
Umweltauswirkungen
Die siebente Veröffentlichung dieses Jahr, die ich zu diesem Thema gefunden habe, war ein hoch interessanter Übersichtsartikel über verschiedene Strahlungen und deren Auswirkungen auf Insekten (9).
Jedem Interessierten rate ich, diesen Artikel mal anzuschauen. Es wird einsichtlich in Diagrammen erklärt, was es überhaupt für Strahlung gibt und wie sich sich auswirken.
Was bei dem Artikel deutlich raus kommt:
Die Lichtverschmutzung ist unser größtes Problem!
Zusammenfassung
Es gibt entgegen vielen Behauptungen einen wissenschaftlichen Konsens in Sachen Mobilfunk. Von daher hat die Neue Zürcher Zeitung (NZZ) natürlich Recht. Ich empfehle dringen allen, die sich erst mal einen Überblick auf Deutsch verschaffen wollen die Lektüre des hervorragend Artikels von Andreas Hirstein am 27.04.2019 in der NZZ

(1) Headache, tinnitus and hearing loss in the international Cohort Study of Mobile Phone Use and Health (COSMOS) in Sweden and Finland (July 2019); International Journal of Epidemiology; Anssi Auvinen et al.; https://doi.org/10.1093/ije/dyz127
(2) Uncertainty Analysis of Mobile Phone Use and Its Effect on Cognitive Function: The Application of Monte Carlo Simulation in a Cohort of Australian Primary School Children (July 2019);
Int. J. Environ. Res. Public Health 2019, 16(13), 2428; https://doi.org/10.3390/ijerph16132428
(3) Radio Frequency Electromagnetic Fields Exposure Assessment in Indoor Environments: A Review (März 2019)
Int. J. Environ. Res. Public Health 2019, 16(6), 955; https://doi.org/10.3390/ijerph16060955
(4) Brain and Salivary Gland Tumors and Mobile Phone Use: Evaluating the Evidence from Various Epidemiological Study Designs. (April 2019); Annual Review of Public Health Volume 40, 2019 Röösli, pp 221-238;
https://www.annualreviews.org/doi/full/10.1146/annurev-publhealth-040218-044037
(5) Mobile phone use and incidence of brain tumour histological types, grading or anatomical location: a population-based ecological study. (Dezember 2018); Public health; Ken Karipidis et al;
https://bmjopen.bmj.com/content/8/12/e024489.long
(6) Mobile Phone Base Station Tower Settings Adjacent to School Buildings: Impact on Students’ Cognitive Health (Januar 2019); American Journal of Men’s Health; Sultan Ayoub Meo et al;
https://journals.sagepub.com/doi/full/10.1177/1557988318816914
(7) Effects of Electromagnetic Radiation of Mobile Phones on the Central Nervous System (2003); Bioelectromagnetics 24:49 ^ 62
https://onlinelibrary.wiley.com/doi/abs/10.1002/bem.10068
(8) Mobile phones, mobile phone base stations and cancer: a review (2005); International Journal of Radiation Biology https://www.tandfonline.com/doi/abs/10.1080/09553000500091097
(9) Risk to pollinators from anthropogenic electro-magnetic radiation (EMR): Evidence and knowledge gaps; Science of The Total; Volume 695, 10 December 2019; Environmenthttps://doi.org/10.1016/j.scitotenv.2019.133833

Text und Abbildungen dürfen unter den Bedingungen der Creative Commons Attribution Share-Alike license (CC-BY-SA) gerne ganz oder teilweise kopiert und zitiert werden.